Способы взятия крови у крыс
У крыс небольшое количество крови удается взять из ушных раковин. Для этого помощник левой рукой фиксирует крысу, крепко зажимая конечности, а большим и указательным пальцами натягивает кожу шеи, сдавливая сосуды этой области и создавая застой крови и гиперемию ушных раковин. Из ушной раковины возможны повторные заборы крови через 3—5 суток. Содержание эритроцитов, лейкоцитов и лейкоцитарная формула крови при повторных взятиях крови из ушной раковины крыс остается без изменений, в то время как при повторных взятиях крови после ампутации кончика хвоста из-за возникающей воспалительной реакции увеличивается число лейкоцитов. Для забора крови из бедренной или яремной вен крыс подвергают наркозу.
Для получения больших количеств крови прибегают у взрослых крыс к пункции хвостовой вены. Хвост обогревают теплой водой, дезинфицируют; вену сдавливают у корня хвоста, вводят в сосуд иглу и шприцем отсасывают кровь. Нередко ДЛЯ взятия крови обрезают кончик хвоста, после чего собирают кровь, вытекающую из раны. Из кончика хвоста удается получить значительное количество крови, вакуумно отсасывая ее. Однако, чтобы взять кровь из вен хвоста, не обязательно отрезать его кончик. Для этого достаточно острой бритвой сделать надрез кончика хвоста наискось, по спирали. Такая рана менее травматична, она быстро заживает.
Кровь из бедренной вены берут под наркозом. Вену отпрепаровывают, вскрывают. Из вскрытого сосуда кровь накопляется в ране. После взятия крови рану тампонируют и зашивают.
Весьма удобно брать кровь у крыс, а также у других мелких лабораторных животных из ретробульбарного венозного сплетения при помощи пастеровской микропипетки (кончик пипетки должен быть слегка заточенным и иметь в диаметре не более 1 мм). Для этого наркотизированную крысу захватывают левой рукой за кожу шеи большим и указательным пальцами, а другими пальцами надежно удерживают за кожу спины. Микропипетку берут в правую руку. Концом пипетки пробуравливающими движениями прокалывают конъюнктиву внутреннего угла глаза и проводят ее на глубину 1—2 мм за глазное яблоко, где находится венозное сплетение. При правильном введении в капилляр микропипетки из ретроорбитального сплетения самотеком поступает кровь. При необходимости взять, большое количество крови следует натянуть кожу в области шеи, чтобы сдавить яремные вены и создать венозный застой, т.е. повысить венозное давление в ретроорбитальном венозном сплетении (рис. 78). При взятии крови из венозного сплетения глазницы необходимо следить, чтобы в пипетку не попала слезная жидкость. Описанный способ прост, позволяет брать кровь при хронических наблюдениях. Редко возникают осложнения в виде повреждения глаза и его слепоты.
Источник
Динамика стресс-активации при повторении стрессогенного воздействия в эксперименте
Цель исследования — анализ динамики стресс-специфичных показателей функционального состояния организма, а именно уровня катехоламинов в плазме крови и спектральных показателей вариабельности сердечного ритма (ВСР), при повторяющемся иммобилизационном стрессе для определения эффектов габитуации.
Материалы и методы. Перед началом эксперимента крыс (Long-Evans, n=6) катетеризировали в яремную вену. Для иммобилизации животных мягко фиксировали в специальном гамаке по 30 мин ежедневно в течение пяти дней. В процессе иммобилизации, а также в течение 30 мин до и 60 мин после иммобилизации в условиях свободного поведения в домашней клетке у крыс проводили неинвазивную регистрацию ЭКГ для последующего анализа ВСР на основе спектральных показателей ритмограмм. В первый, третий и пятый экспериментальные дни у крыс брали пробы крови за 30 мин до (1-й этап), через 5 мин после начала иммобилизации (2-й этап), в конце иммобилизации (3-й этап) и через 30 мин после окончания иммобилизационного периода (4-й этап) для последующего анализа концентрации катехоламинов в плазме крови.
Результаты. Эффекты габитуации к иммобилизации выявлены в динамике уровня катехоламинов в плазме крови, ЧСС и ВСР. Концентрация адреналина и норадреналина увеличивалась статистически значимо в период иммобилизации в первый экспериментальный день и не имела достоверных изменений в последующие дни. ЧСС статистически значимо снижалась на 3-м и 4-м этапах эксперимента (свободное поведение в домашней клетке после иммобилизации) в пятый день и оставалась на исходно высоком уровне в первый и третий дни эксперимента. Общая мощность спектра ВСР статистически значимо уменьшалась в период иммобилизации во все экспериментальные дни. В первый день ВСР оставалась редуцированной до 4-го этапа эксперимента. В третий и пятый дни общая мощность спектра ВСР восстанавливалась после иммобилизации до исходного уровня к 4-му этапу эксперимента.
Заключение. Пятидневное повторение иммобилизационного стресса животных в специальном гамаке, разработанном для их фиксации в электрофизиологических экспериментах, сопровождается габитуацией, что демонстрируют данные динамики уровня катехоламинов в крови и ВСР. Полученные данные показывают, что эффекты габитуации проявляются не только в снижении выраженности стресс-активации, но и в сокращении ее по времени.
Интерес к хроническому стрессу обусловлен увеличением количества заболеваний (особенно сердечно-сосудистых), являющихся его следствием [1]. Исследования хронического стресса в основном акцентируются на аспектах динамики состояния индивида и выяснении его маркеров для повышения эффективности диагностики [2]. Для этого разрабатываются экспериментальные процедуры формирования хронического стресса у индивидов [1]. В таких экспериментальных процедурах (например, модель непредсказуемого хронического неострого стресса [3]) большое внимание уделяется избеганию эффектов габитуации — привыкания индивида к первоначально стрессогенному воздействию. Однако феноменология габитуации и ее динамики мало исследованы, как и в целом факторы, определяющие процессы, разворачивающиеся при повторяющемся стрессе [4].
Эффекты габитуации (или, наоборот, сенситизации) при повторении стресса обычно исследуются для тестирования моделей стрессогенных воздействий на возможность их неоднократного использования в экспериментах [5]. На уровне биохимических показателей не выявлено согласованности эффектов в отношении габитуации к повторяющемуся стресс-фактору [4]. Так, при повторении распространенной модели социального стресса (trier social stress test) показано, что эффекты габитуации проявляются в динамике концентрации кортизола (показатель активации гипоталамо-гипофизарной оси) и альфа-амилазы (показатель активации α-адренорецепторов) в слюне и не наблюдаются в динамике концентрации интерлейкина-6 (показатель выраженности воспалительных процессов). На физиологическом уровне некоторые характеристики вариабельности сердечного ритма (ВСР) — среднее значение и дисперсия R–R-интервалов — предлагаются как возможный способ оценки эффектов габитуации при повторяющемся когнитивном стрессе [6]. Кроме того, динамика этих показателей была согласована с поведенческими эффектами габитуации. Выраженность эффектов габитуации, оцениваемая по изменениям частоты сердечных сокращений (ЧСС), систолическому и диастолическому давлению, прямо коррелировала с показателями открытости новому опыту, оцениваемыми на основе опросника [7].
Имеющиеся данные обусловливают дальнейшее исследование динамики и механизмов габитуации к повторяющимся стрессогенным воздействиям как простейшей формы научения, выражающейся в снижении проявления симптомов, специфичных стрессу.
Кроме прикладного аспекта исследования габитуации при повторении стресса имеют и методическую актуальность. Как правило, экспериментальные протоколы с животными включают первоначальный этап привыкания (к обстановке или необходимому, но не исследуемому воздействию) для устранения эффектов новизны и стресса в динамике интересующих переменных. Более контактные процедуры (например, фиксация животного в установке) требуют более длительного периода привыкания. Дополнительное использование в эксперименте способов контроля состояния животного позволит надежно исключить возможность стрессогенного воздействия выбранной процедуры. Отсутствие такой возможности требует проведения предварительной пилотажной серии экспериментов с целью определения этапа, на котором у большинства или всех животных отсутствует стресс при реализации экспериментального протокола. В этом направлении наиболее интересным является исследование габитуации при повторяющемся иммобилизационном стрессе, так как многие эксперименты электрофизиологических исследований включают разной степени ограничения подвижности животного нефармакологическими способами.
Цель исследования — определение динамики функционального состояния индивида (по показателям, специфичным для состояния стресса) при повторении стрессогенного воздействия в модели иммобилизационного стресса.
Материалы и методы
Исследование проводилось на половозрелых самцах крыс линии Long-Evans (средняя масса животных составляла 300 г, средний возраст — 19 мес (мин=6, макс=24)). Катетеризировали яремную вену у 39 крыс, однако в связи с техническими проблемами (непроходимость катетера) полную экспериментальную серию удалось выполнить только на 6 животных.
Крысы содержались в стандартных условиях вивария в одиночных клетках и имели свободный доступ к воде и пище. Все манипуляции с ними выполняли в соответствии с Приказом №199н «Об утверждении Правил надлежащей лабораторной практики» (Россия, 2016) и «Международными рекомендациями (этический кодекс) по проведению медико-биологических исследований с использованием животных» (CIOMS и ICLAS, 2012), при этом неукоснительно соблюдались этические принципы, установленные Европейской конвенцией по защите позвоночных животных, используемых для экспериментальных и других научных целей (Страсбург, 2006). На проведение экспериментальных исследований на животных получено разрешение этического комитета Института психологии РАН.
Операция по установке катетера в яремную вену. Катетеры состояли из силиконовой трубки (короткой, 2,5 см) и полиэтиленовой трубки (длинной, 6 см). Стерильные катетеры промывались раствором медицинского спирта 96%. Для наркоза животным вводили раствор Рометара (10 мг/кг) и Золетила (25 мг/кг) внутримышечно. В операционном поле выстригали волосяной покров, брили и обрабатывали антисептиком. Разрез кожных покровов производили по косой линии, соединяющей верхнюю треть грудины и внутреннюю треть левой или правой ключицы таким образом, чтобы 2/3 разреза находились ниже ключицы. Длина разреза составляла 1–1,5 см. Далее рассекали поверхностную фасцию и яремную вену отпрепаровывали тупым способом. Затем на яремную вену ставили лигатуры: ростральнее — постоянную, каудальнее — временную. Между ними выполняли надрез внешней стенки яремной вены. Полученное отверстие расширяли браншами микрохирургического пинцета и вводили силиконовую часть катетера в каудальном направлении, ослабляя каудальную лигатуру, на глубину 2–3 см. Введенный катетер проверяли на возможность забора крови, заполняли гепаринизированным физиологическим раствором (500 ЕД/мл, 80 мкл), закрывали пробкой и фиксировали лигатурами на вене. Затем подкожно выводили его через надрез (3–4 мм) между лопатками, после чего ушивали раны.
Контроль проходимости катетера проводили ежедневно в течение недели взятием пробы крови.
Протокол эксперимента. Через неделю после операции по установке катетера выполняли пятидневную экспериментальную серию с ежедневными сессиями, каждая из которых включала 4 этапа (рис. 1). На 1-м этапе (30 мин) крыса находилась в домашней клетке в условиях свободного поведения. На 2-м этапе (30 мин) крыса фиксировалась в специальном гамаке для иммобилизации. На 3-м и 4-м этапах (по 30 мин) крыса снова находилась в домашней клетке в условиях свободного поведения.
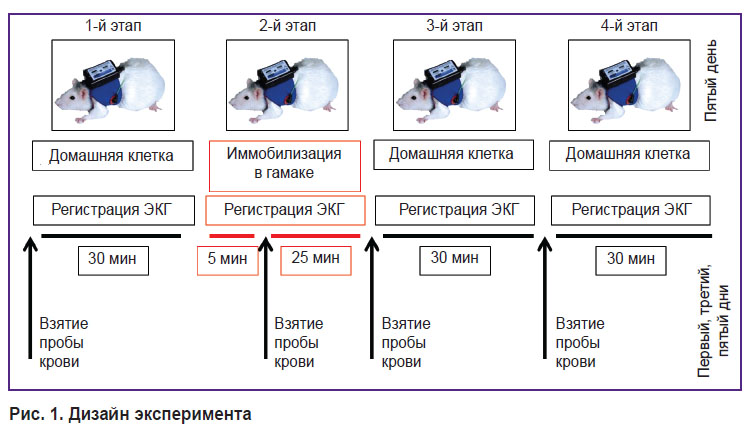 | Рис. 1. Дизайн эксперимента |
На каждом этапе проводили неинвазивную регистрацию ЭКГ.
В первый, третий и пятый дни эксперимента перед 1, 3, 4-м этапами и на 5-й минуте 2-го этапа производили забор крови через катетер (1 мл). Взятую пробу крови центрифугировали для отделения фракции плазмы и хранили при t=18°C.
Регистрация сердечного ритма. Для неинвазивной биполярной регистрации ЭКГ у крыс использовали систему «Физиобелт» (МПК «Открытая наука», Россия), которая включает манжетку для постановки электродов на тело животного и крепления прибора со встроенными усилителем, аналого-цифровым преобразователем и передатчиком. Прибор имеет размеры 35×55×15 мм и массу 40 г. Частота опроса — 500 Гц. При регистрации использовали программное обеспечение Physiobelt (ООО «Нейроботикс», Россия).
Определение концентрации катехоламинов в плазме крови. Оценку уровня катехоламинов в плазме крови проводили на изократическом хроматографе Gilson 407 (Gilson, Франция) с использованием электрохимического детектора LC-4C (BAS, США) по протоколу М.Х. Каримовой и соавт. [8]. Все реактивы были степени Х.Ч., analytical grade.
В образцах плазмы крови определяли концентрацию адреналина и норадреналина. Эти вещества были выбраны для анализа, поскольку возрастание их концентрации является маркером I стадии острого стресса (стадии активации симпатоадреналовой системы [9]).
Анализ вариабельности сердечного ритма. В ЭКГ крыс выделяли последовательности R–R-интервалов (временных интервалов между R-зубцами в соседних QRST-комплексах кардиограммы) пороговым методом в программе Spike 2.5. Для выделения ритмограмм выбирались 100-секундные интервалы ЭКГ без артефактов. Полученные ритмограммы анализировали с построением спектров периодограммой Уэлча, предварительно интерполировав их с шагом 50 мс. Для этого использовали программу Rhythm Service 1.2 (ООО «Фотон-тест», Россия). В результате для каждой последовательности R–R-интервалов вычисляли ЧСС (в минуту) — среднюю частоту сердечных сокращений; TP (мс 2 ) — суммарную мощность спектра ВСР; LF (мс 2 ) — мощность спектра в области низких частот (0,04–0,15 Гц); HF (мс 2 ) — мощность спектра в области высоких частот (0,15–0,60 Гц); LF/HF — отношение мощностей спектра в области низких и высоких частот (индекс вегетативного баланса). Все описанные показатели рассчитывались для каждого этапа эксперимента как средние значений для всех выделенных ритмограмм длительностью 100 с в этот период.
Спектральные показатели были выбраны для характеристики ВСР, так как предыдущие работы [10, 11] показали, что снижение общей мощности спектра ВСР и возрастание индекса вегетативного баланса специфичны для начала острого стресса.
Статистические процедуры. Для первого, третьего и пятого экспериментальных дней динамику показателей уровня катехоламинов в плазме крови и спектральных показателей ВСР в течение эксперимента (от 1-го до 4-го этапа) проверяли на наличие достоверных изменений, используя критерий Фридмана [12]. Парные сравнения переменных между этапами эксперимента и между экспериментальными днями проводили с использованием критерия Вилкоксона [13]. Изменения считались статистически значимыми при р≤0,05.
Результаты и обсуждение
Анализ динамики концентрации адреналина показал наличие статистически значимых изменений ( 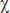 2 (n=6; df=3)=14,8; p=0,002, критерий Фридмана) в первый день экспериментальной серии. Отмечено статистически значимое возрастание адреналина на 2-м этапе эксперимента (при иммобилизации животного) и последующее снижение к 3-му, а затем — к 4-му этапу (свободное поведение в домашней клетке) (р≤0,05, критерий Вилкоксона). Аналогичные изменения наблюдались и в динамике норадреналина (
2 (n=6; df=3)=14,8; p=0,002, критерий Фридмана) в первый день экспериментальной серии. Отмечено статистически значимое возрастание адреналина на 2-м этапе эксперимента (при иммобилизации животного) и последующее снижение к 3-му, а затем — к 4-му этапу (свободное поведение в домашней клетке) (р≤0,05, критерий Вилкоксона). Аналогичные изменения наблюдались и в динамике норадреналина ( 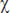 2 (n=6; df= 3)=8,8; p=0,03, критерий Фридмана).
2 (n=6; df= 3)=8,8; p=0,03, критерий Фридмана).
На третий день эксперимента статистически значимые изменения отсутствовали в динамике как адреналина ( 2 =3,8; p=0,28, критерий Фридмана), так и норадреналина ( 2 =1,0; p=0,80, критерий Фридмана). Аналогично и в пятый день эксперимента отсутствовали достоверные изменения в динамике адреналина ( 2 =6,0; p=0,11, критерий Фридмана) и норадреналина ( 2 =2,4; p=0,49, критерий Фридмана).
Значения адреналина и ноадреналина были статистически значимо выше на 2-м этапе эксперимента в первый день, чем на 2-м этапе в третий и пятый дни (р≤0,05, критерий Вилкоксона — для обеих переменных). На 1, 3 и 4-м этапах значения адреналина и норадреналина не отличались статистически значимо в первый, третий и пятый экспериментальные дни.
Подробнее динамика адреналина и норадреналина в течение всех дней эксперимента показана на рис. 2.
Рис. 2. Динамика концентрации адреналина и норадреналина (нг/мл) в плазме крови крыс в эксперименте с иммобилизацией (значения медиан и процентилей распределения):
1 — в начале 1-го этапа эксперимента (30 мин до иммобилизации); 2 — на 5-й минуте второго этапа (иммобилизация в гамаке); 3 — в начале 3-го этапа (сразу по окончании иммобилизации); 4 — в начале 4-го этапа эксперимента (через 30 мин после иммобилизации) в первый, третий и пятый дни экспериментальной серии; * — p≤0,05, критерий Вилкоксона; # — p≤0,05, критерий Фридмана
Таким образом, по данным концентрации катехоламинов в плазме крови, в первый день у животных при иммобилизации в экспериментальном гамаке наблюдается стресс-активация, которая проявляется в статистически значимом возрастании уровня адреналина и норадреналина и возвращении к исходному уровню через 30 мин после окончания иммобилизации. На третий и пятый дни в динамике концентрации катехоламинов зафиксированы эффекты габитуации к иммобилизации, так как отсутствуют достоверные изменения адреналина и норадреналина между этапами эксперимента, а также установлено их статистически значимое снижение при иммобилизации по сравнению с первым экспериментальным днем. Можно предположить, что габитуация к иммобилизации в данном случае проявляется не в форме редукции стресс-активации, а в сокращении временного периода, в который она разворачивается. В таком случае, если стресс-активация запускается и заканчивается в течение первых 5 мин иммобилизации, во второй пробе плазмы крови (которая проводилась через 5 мин после начала 2-го этапа эксперимента) фиксируется только исходный уровень адреналина и норадреналина.
В динамике ЧСС статистически значимых изменений не наблюдалось в первый ( 2 =4,00; p=0,261, критерий Фридмана) и третий ( 2 =3,4; p=0,334, критерий Фридмана) дни эксперимента. При этом были выявлены достоверные изменения в пятый день ( 2 =11,6; p=0,009, критерий Фридмана): наблюдалось статистически значимое снижение ЧСС от 2-го этапа эксперимента (период иммобилизации) к 3-му и 4-му этапам (свободное поведение в домашней клетке). Подробнее динамика ЧСС в течение всех дней эксперимента показана на рис. 3.
Рис. 3. Динамика ЧСС, общей мощности спектра ВСР (TP) и индекса вегетативного баланса (LF/HF) (значения медиан и процентилей распределения) у крыс в эксперименте с иммобилизацией:
1 — во время 1-го этапа эксперимента (30 мин свободного поведения в домашней клетке); 2 — во время 2-го этапа (30 мин иммобилизации в гамаке), 3 — во время 3-го этапа (30 мин свободного поведения в домашней клетке); 4 — во время 4-го этапа (30 мин свободного поведения в домашней клетке) в первый, третий и пятый дни экспериментальной серии; * — p≤0,05, критерий Вилкоксона; # — p≤0,05, критерий Фридмана